투푸수아라
백악기 잡식 생물 종류
Tupuxuara longicristatus
학명: "투피(Tupi)족 신화의 '친숙한 영혼(familiar spirit)' + longicristatus = 라틴어 '긴 볏'"
신체 특징
발견
서식지

투푸수아라(Tupuxuara Kellner & Campos, 1988)는 백악기 전기 알비안절(약 1억 1200만 년 전)에 현재의 브라질 북동부 아라리페 분지(Araripe Basin)에 서식했던 대형 익룡이다. 익룡목(Pterosauria) 익룡아목(Pterodactyloidea)의 아즈다르코이데아(Azhdarchoidea) 상과에 속하며, 탈라소드로미과(Thalassodromidae)의 대표적인 구성원이다. 속명 'Tupuxuara'는 브라질 원주민 투피(Tupi)족 신화에 등장하는 '친숙한 영혼(familiar spirit)'을 뜻하며, 모식종의 종소명 longicristatus는 라틴어로 '긴 볏'을 의미한다.
투푸수아라의 가장 두드러진 특징은 주둥이부터 머리 뒤쪽까지 뻗어 있는 거대한 골질 볏(bony sagittal crest)이다. 이빨이 없는(edentulous) 긴 부리를 가졌으며, Gregory S. Paul(2022)의 추정에 따르면 날개폭 약 4.7 m, 전체 체장 약 2 m, 체중 약 25 kg으로 비교적 큰 편이었다. 가장 큰 표본(T. leonardii 참조 표본)의 두개골 길이는 약 130 cm에 달하며, 이에 기초한 날개폭 추정치는 약 5.5 m에 이른다. 현재 유효종으로 인정되는 종은 T. longicristatus(모식종)와 T. leonardii 두 종이며, 2009년에 기재된 T. deliradamus는 2013년 이후 의문명(nomen dubium)으로 취급되는 추세이다.
초기 연구에서는 투푸수아라의 식성이 어식(piscivorous)으로 추정되었으나, 아즈다르코이데아 계통 내 위치와 형태학적 특징을 고려하면 육상성 잡식 또는 육식 동물이었을 가능성이 더 높다는 것이 최근의 주류 견해이다. 공룡이 아닌 익룡으로, 공룡과는 독립적인 진화 계통에 속하는 비행 파충류이다.
개요
이름과 어원
속명 Tupuxuara는 브라질 원주민 투피(Tupi)족 신화에 등장하는 친숙한 영혼(familiar spirit)에서 유래했다. Paleofile.com에 의하면 투피 문화에서 남성 친숙한 영혼(가정의 시종적 존재)을 가리키는 말이다. 모식종 T. longicristatus의 종소명은 라틴어 longus(긴)와 cristatus(볏이 있는)의 합성어로, 이 익룡의 길게 뒤로 젖혀진 두개골 볏을 묘사한 것이다(Kellner & Campos, 1988). 두 번째 종 T. leonardii의 종소명은 이탈리아 출신 고생물학자이자 지질학자인 Giuseppe Leonardi를 기리기 위해 명명되었다(Kellner & Campos, 1994).
분류 상태
현재 유효하게 인정되는 종은 T. longicristatus(1988)와 T. leonardii(1994) 두 종이다. 2009년 Mark Witton이 기재한 세 번째 종 T. deliradamus는 Kellner(2013)에 의해 의문명으로 판정되었고, Cerqueira et al.(2023)의 계통분석에서는 Tupuxuara가 아닌 Caupedactylus ybaka의 자매군으로 회수되어 사실상 속 자체에서 제외되었다. 따라서 현재 학계에서는 투푸수아라를 두 종으로 구성된 속으로 취급하는 것이 지배적이다.
한 줄 요약
투푸수아라는 브라질 로무알도층에서 산출된 대형 무치(toothless) 탈라소드로미드 익룡으로, 거대한 골질 두개골 볏과 상대적으로 가벼운 몸체가 특징이다.
시대, 층서, 산출 환경
시대 범위
투푸수아라의 화석은 브라질 아라리페 분지(Araripe Basin)의 로무알도층(Romualdo Formation, Santana Group)에서 산출된다. 로무알도층의 연대는 초기 백악기 알비안절에 해당하며, 약 1억 1100만~1억 800만 년 전(111~108 Ma)으로 추정된다. 대부분의 연구자들은 압티안-알비안 경계(약 112 Ma) 부근에 이 지층이 위치한다는 데 동의하지만, 일부 연구에서는 세노마니안(Cenomanian) 가능성도 완전히 배제하지 않고 있다(Martill et al., 2007).
지층과 암상
로무알도층은 산타나 그룹(Santana Group)에 속하며, 이전에는 산타나층(Santana Formation)의 로무알도 부층원(Romualdo Member)으로 불렸다. 주요 암상은 이암(mudstone)이며, 석회암(limestone)과 셰일(shale)이 수반된다. 특히 이 지층은 탄산염 교결체(carbonate concretions)가 특징적이며, 이 교결체 내부에서 연부조직까지 보존된 뛰어난 화석이 산출되어 콘서바트-라거슈테테(Konservat-Lagerstätte)로 지정되어 있다.
퇴적 환경과 고환경
로무알도층은 열개 분지(rift basin) 내 호수-석호 환경에 원시 대서양(proto-Atlantic)의 얕은 해수가 유입되는 환경에서 퇴적되었다. 당시 남대서양이 열리면서 좁고 얕은 해로(海路)가 형성되었으며, 투푸수아라는 이 연안-석호(lagoonal/restricted shallow marine) 환경에서 서식했을 것으로 추정된다. 고위도·고경도(paleocoordinates) 분석에 따르면 당시 이 지역은 약 남위 12.2°, 서경 10.7°에 위치해 있었다(Romualdo Formation Wikipedia paleocoordinates).
표본 및 진단 형질
주요 표본
| 종 | 표본 번호 | 구성 부위 | 기관/비고 |
|---|---|---|---|
| T. longicristatus (모식종) | MN 6591-V (홀로타입) | 부분적 주둥이(snout)와 일부 날개 골편(wing metacarpal, 1st wing phalanx) | 브라질 국립박물관(Museu Nacional) |
| T. leonardii | MN 6592-V (홀로타입) | 단편적 두개골, 더 둥근 볏 | 브라질 국립박물관 |
| T. leonardii (참조 표본) | IMCF 1052 | 거의 완전한 두개골 (약 103 cm 길이) | 이와키시 석탄화석관(일본) |
| T. leonardii (대형 표본) | 비공식 | 두개골 길이 약 130 cm, 날개폭 5.5 m 추정 | 개인/기관 미상 |
| T. deliradamus (의문명) | SMNK PAL 6410 (홀로타입) | 두개골 | SMNK(독일) |
| T. deliradamus (부모식) | KPMNH DL 84 | 두개골 | KPMNH(일본) |
진단 형질
투푸수아라를 다른 아즈다르코이드 익룡과 구분짓는 주요 진단 형질은 다음과 같다. 첫째, 주둥이에서 머리 뒤쪽까지 연장되는 커다란 골질 정중선 볏(bony midline crest)을 가진다. 둘째, 이빨이 전혀 없는 무치성(edentulous) 부리를 가진다. 셋째, 비전안와공(nasoantorbital fenestra) 아래에 강하게 볼록한 상악 구개판(convex maxillary palatal plates)이 있다(Witton, 2009). T. longicristatus는 뒤로 길게 젖혀진(backswept) 볏을, T. leonardii는 상대적으로 더 둥근(rounded) 볏을 가진다는 점에서 두 종이 구분된다.
표본의 한계
홀로타입 MN 6591-V는 주둥이와 일부 날개 골편만으로 구성되어 있어 완전한 골격 복원에는 한계가 있다. T. leonardii의 경우 IMCF 1052 등 비교적 완전한 두개골 자료가 있으나, 두개골 이후(postcranial) 골격의 연관 자료는 여전히 부족하다. 따라서 체장이나 체중 추정은 다른 탈라소드로미드나 아즈다르코이드 익룡과의 비교에 의존하는 부분이 크다.
형태와 기능
체형과 크기
Gregory S. Paul(2022)은 The Princeton Field Guide to Pterosaurs에서 투푸수아라의 날개폭을 약 4.7 m, 전체 체장을 약 2 m, 체중을 약 25 kg으로 추정했다. Paul은 투푸수아라가 체중에 비해 상당히 긴 날개를 가졌다고 언급했다. 한편, 가장 큰 T. leonardii 표본(두개골 길이 약 130 cm)에 기초한 추정 날개폭은 약 5.5 m에 달하며, 이는 속 내에서도 상당한 개체 크기 변이가 존재했음을 시사한다.
두개골과 볏
투푸수아라의 가장 눈에 띄는 형태적 특징은 두개골의 거대한 골질 볏이다. 이 볏은 주둥이 앞부분에서 기원하여 두개골 뒤쪽까지 연장되며, 주로 골질(bone)로 이루어져 있다. Martill & Naish(2006)는 어린 개체(subadult)에서는 볏이 아직 완전히 발달하지 않았음을 보고하여, 볏이 성적 성숙의 지표(sexual maturity marker)로 기능했을 가능성을 제시했다. 두 종 사이의 주요 차이도 볏의 형태에서 나타나는데, T. longicristatus는 길고 삼각형에 가까운 뒤로 젖혀진 볏을, T. leonardii는 더 둥글고 부채꼴에 가까운 볏을 가진다.
부리와 치열
투푸수아라는 완전한 무치성(toothless) 익룡으로, 이빨이 전혀 없는 각질 부리를 가졌을 것으로 추정된다. 이는 같은 과의 Thalassodromeus와 공유하는 특징이며, 타페자라과(Tapejaridae)의 다른 구성원들과도 유사하다. 다만 투푸수아라의 부리는 Thalassodromeus보다 덜 견고한(less heavily built) 구조였다(Witton, 2013).
날개와 비행
익룡의 날개는 네 번째 손가락이 극도로 길어져 형성된 비막(wing membrane)으로 구성된다. 투푸수아라의 경우 체중에 비해 날개폭이 상당히 크며(Paul, 2022), 이는 효율적인 활공(soaring)에 적합한 형태이다. 익룡의 골격은 속이 빈(pneumatic) 중공 구조로 매우 가벼우면서도 내부 보강 구조를 통해 충분한 강도를 유지했다.
식성 및 생태
식성 논쟁
투푸수아라의 식성은 상당한 논쟁의 대상이 되어왔다. 초기 연구에서는 로무알도층의 해양/석호 환경과 긴 부리 형태에 기초하여 어식(piscivory)으로 추정했다. 일부 이례적인 가설에서는 과일을 먹는 식성(frugivory)까지 제안되었다. 그러나 최근의 주류 견해는 투푸수아라의 아즈다르코이드 계통상 위치를 고려하면 육상성 잡식(omnivore) 또는 육식(carnivore)이었을 가능성이 높다는 것이다(Wikipedia, Tupuxuara; Witton, 2013; Pteros.com). 근연종 Thalassodromeus가 더 큰 먹이에 특화된 것과 달리, 투푸수아라 두 종은 그러한 특화가 관찰되지 않는다.
활동 패턴
Schmitz & Motani(2011)는 공막환(scleral ring)과 안와(orbit) 형태를 비교 분석한 연구에서, 투푸수아라의 눈 구조가 현생 주행성(diurnal) 조류 및 파충류와 유사하다는 결과를 보고했다. 이는 투푸수아라가 주로 낮에 활동했음을 시사한다.
생태적 지위
투푸수아라는 로무알도층의 풍부한 동물상과 공존했다. 같은 지층에서 산출되는 익룡으로는 대형 어식성 Tropeognathus, Anhanguera 속의 여러 종, 그리고 같은 탈라소드로미과의 Thalassodromeus sethi, Kariridraco dianae, 타페자라과의 Tapejara wellnhoferi 등이 있다. 육상에는 스피노사우루스과(Irritator), 콤프소그나투스과(Mirischia) 등의 수각류 공룡도 서식했다. 투푸수아라는 이들 어식성 익룡들과는 다른 생태적 지위를 점했을 가능성이 높으며, 육상 환경에서 소형 동물이나 다양한 먹이를 섭식하는 범식자(generalist feeder)였을 것으로 추정된다.
분포와 고지리
산지 분포
투푸수아라의 모든 확실한 표본은 브라질 북동부 아라리페 분지의 로무알도층에서 산출되었다. 이 지역은 현재 세아라(Ceará), 페르남부코(Pernambuco), 피아우이(Piauí) 세 주의 경계에 위치하며, 아라리페 고원(Chapada do Araripe) 기저부에 해당한다.
고지리 해석
로무알도층의 고좌표(paleocoordinates)는 약 남위 12.2°, 서경 10.7°로 추정된다. 백악기 알비안절에 이 지역은 남대서양이 열리기 시작하는 좁고 얕은 해협(seaway) 인근에 위치해 있었으며, 아열대 기후의 해안-석호 환경이었을 것이다. 탈라소드로미과의 모든 확실한 구성원(Tupuxuara, Thalassodromeus, Kariridraco)이 동일한 로무알도층에서만 알려져 있어, 이 그룹이 백악기 전기 브라질 지역에 국한된 분포를 보였을 가능성이 있다.
계통과 분류 논쟁
분류사
투푸수아라의 분류학적 위치는 익룡 계통분류에서 가장 논쟁적인 주제 중 하나이다. Kellner는 원기재(1988) 이래 투푸수아라를 타페자라과(Tapejaridae) 내 탈라소드로미아(Thalassodrominae) 아과에 배치해왔다(Kellner & Campos, 2007). 이 체계에서 투푸수아라는 Thalassodromeus와 함께 타페자라과 내에서 골질 볏을 공유하는 클레이드를 형성한다.
그러나 Unwin(2003) 이래 다수의 연구(Martill & Naish, 2006; Witton, 2009; Longrich et al., 2018; Andres, 2021)는 투푸수아라가 타페자라과보다 아즈다르키과(Azhdarchidae)에 더 가깝다는 결과를 보고했다. 이 체계에서는 투푸수아라와 Thalassodromeus를 포함하는 그룹이 독립적인 과(Family) 수준의 탈라소드로미과(Thalassodromidae, Witton 2009)로 격상되며, 둥다리프테루스과(Dsungaripteridae)와 함께 네오아즈다르키아(Neoazhdarchia)를 구성한다.
최근 합의 동향
Pêgas et al.(2023)은 두 체계 간의 명칭 혼란을 해소하기 위해, 탈라소드로미과(Thalassodromidae)라는 과(Family) 수준의 명칭을 일관되게 사용할 것을 제안했다. 이 연구에서 탈라소드로미과는 T. longicristatus, T. leonardii, Thalassodromeus, Kariridraco를 포함하며, 타페자라과와 함께 타페자라형태군(Tapejaromorpha)을 구성하는 것으로 회수되었다. 한편, Andres(2021)의 체계에서는 탈라소드로미과가 네오아즈다르키아 내 둥다리프테루스형태군(Dsungaripteromorpha)에 포함되어 타페자라과와는 더 먼 관계로 회수된다.
주요 분류 비교
| 계통 모형 | 탈라소드로미과 위치 | 근연 그룹 | 대표 연구 |
|---|---|---|---|
| Kellner 모형 (Tapejaromorpha) | 타페자라형태군 내 독립 과 | 타페자라과(Tapejaridae) 자매군 | Pegas et al. 2023 |
| Unwin 모형 (Neoazhdarchia) | 네오아즈다르키아 내 독립 과 | 아즈다르키과 + 둥다리프테루스과 | Andres 2021; Longrich et al. 2018 |
복원과 불확실성
확정된 사항
투푸수아라가 대형 무치성 익룡이었으며, 거대한 골질 볏을 가졌다는 점은 다수의 표본에서 확인된 사실이다. 아즈다르코이데아 상과 내에서의 소속도 모든 계통분석에서 일관되게 지지된다. 로무알도층(알비안, 약 112 Ma)에서의 산출 역시 확정적이다.
유력한 추정
볏이 성적 성숙의 표지(sexual maturity indicator)로 기능했다는 가설은 어린 개체의 미발달 볏이라는 직접적 증거가 있어 유력하다(Martill & Naish, 2006). 주행성(diurnal) 활동 패턴은 공막환 분석에 기초한 유력 추정이다(Schmitz & Motani, 2011).
가설/논쟁 중인 사항
탈라소드로미과가 타페자라과에 가까운지(Kellner 모형) 아즈다르키과에 가까운지(Unwin 모형)는 여전히 해결되지 않은 핵심 논쟁이다. 식성 역시 어식(초기 가설) vs. 육상 잡식/육식(최근 주류)으로 논쟁이 있으며, 직접적인 위내용물 같은 확정적 증거는 아직 보고되지 않았다. 볏의 공기역학적 기능(비행 중 방향타 역할) 역시 제안된 바 있으나, 성적 선택(sexual selection)이나 종 인식(species recognition) 기능이 더 유력시된다.
대중 매체 vs. 학계
투푸수아라는 대중 매체에서 종종 물고기를 사냥하는 해양성 익룡으로 묘사되지만, 최근 학계에서는 육상 활동이 더 주된 생활 방식이었을 것으로 보고 있다. 또한 공룡이 아닌 익룡임을 혼동하는 경우가 흔하며, 익룡은 공룡과는 별개의 분류군(Archosauria 내에서 독립적 계통)이라는 점을 주의해야 한다.
근연종 및 동시대 비교
| 분류군 | 날개폭 | 볏 형태 | 식성 추정 | 시대/지층 |
|---|---|---|---|---|
| Tupuxuara longicristatus | ~4.7 m | 길게 뒤로 젖혀진 골질 볏 | 잡식/육식(추정) | 알비안, 로무알도층 |
| Tupuxuara leonardii | ~5.5 m (대형 개체) | 둥근 부채꼴 골질 볏 | 잡식/육식(추정) | 알비안, 로무알도층 |
| Thalassodromeus sethi | ~4.2~4.5 m | 매우 크고 편평한 골질 볏 (두개골 ~1.42 m) | 대형 먹이 특화(추정) | 알비안, 로무알도층 |
| Kariridraco dianae | 미확정 | 골질 볏 | 미확정 | 알비안, 로무알도층 |
| Tapejara wellnhoferi | ~3.5 m | 골질 + 연조직 볏 | 잡식(추정) | 알비안, 로무알도층 |
투푸수아라는 Thalassodromeus보다 약간 작고, 두개골이 덜 견고하며 볏의 크기도 상대적으로 작다. 반면 같은 지층의 타페자라(Tapejara)보다는 훨씬 크고, 볏이 주로 골질로 이루어져 있다는 점에서 구별된다.
재미있는 사실
FAQ
📚참고문헌
- Kellner, A. W. A. & Campos, D. A. (1988). Sobre um novo pterossauro com crista sagital da Bacia do Araripe, Cretáceo Inferior do Nordeste do Brasil. Anais da Academia Brasileira de Ciências, 60: 459–469.
- Kellner, A. W. A. & Campos, D. A. (1994). A new species of Tupuxuara (Pterosauria, Tapejaridae) from the Early Cretaceous of Brazil. Anais da Academia Brasileira de Ciências, 66: 467–473.
- Martill, D. M. & Naish, D. (2006). Cranial crest development in the Azhdarchoid pterosaur Tupuxuara, with a review of the genus and tapejarid monophyly. Palaeontology, 49(4): 925–941. doi:10.1111/j.1475-4983.2006.00575.x
- Kellner, A. W. A. & Campos, D. A. (2007). Short note on the ingroup relationships of the Tapejaridae (Pterosauria, Pterodactyloidea). Boletim do Museu Nacional: Geologia, 75: 1–14.
- Witton, M. P. (2009). A new species of Tupuxuara (Thalassodromidae, Azhdarchoidea) from the Lower Cretaceous Santana Formation of Brazil, with a note on the nomenclature of Thalassodromidae. Cretaceous Research, 30(5): 1293–1300. doi:10.1016/j.cretres.2009.07.006
- Schmitz, L. & Motani, R. (2011). Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology. Science, 332(6030): 705–708. doi:10.1126/science.1200043
- Kellner, A. W. A. (2013). A new unusual tapejarid (Pterosauria, Pterodactyloidea) from the Early Cretaceous Romualdo Formation, Araripe Basin, Brazil. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 103(3–4): 409–421. doi:10.1017/S1755691013000327
- Witton, M. P. (2013). Pterosaurs: Natural History, Evolution, Anatomy. Princeton University Press. ISBN 0691150613.
- Longrich, N. R., Martill, D. M., Andres, B. & Penny, D. (2018). Late Maastrichtian pterosaurs from North Africa and mass extinction of Pterosauria at the Cretaceous-Paleogene boundary. PLOS Biology, 16(3): e2001663. doi:10.1371/journal.pbio.2001663
- Andres, B. (2021). Phylogenetic systematics of Quetzalcoatlus Lawson 1975 (Pterodactyloidea: Azhdarchoidea). Journal of Vertebrate Paleontology, 41(sup1): 203–217. doi:10.1080/02724634.2020.1801703
- Paul, G. S. (2022). The Princeton Field Guide to Pterosaurs. Princeton University Press. p. 156. doi:10.1515/9780691232218
- Pêgas, R. V., Zhou, X., Jin, X., Wang, K. & Ma, W. (2023). A taxonomic revision of the Sinopterus complex (Pterosauria, Tapejaridae) from the Early Cretaceous Jehol Biota, with the new genus Huaxiadraco. PeerJ, 11: e14829. doi:10.7717/peerj.14829
- Cerqueira, G. M., Müller, R. T. & Pinheiro, F. L. (2023). On the phylogenetic affinities of the tapejarid pterosaur 'Tupuxuara deliradamus' from the Lower Cretaceous of Brazil. Historical Biology, 36(3): 677–682. doi:10.1080/08912963.2023.2180741
- Fara, E., Saraiva, A. Á. F., Campos, D. A., Moreira, J. K. R., de Carvalho Siebra, D. & Kellner, A. W. A. (2005). Controlled excavations in the Romualdo Member of the Santana Formation (Early Cretaceous, Araripe Basin, northeastern Brazil): stratigraphic, palaeoenvironmental and palaeoecological implications. Palaeogeography, Palaeoclimatology, Palaeoecology, 218(1–2): 145–160. doi:10.1016/j.palaeo.2004.12.012
- Pêgas, R. V., Costa, F. R. & Kellner, A. W. A. (2018). New Information on the osteology and a taxonomic revision of the genus Thalassodromeus (Pterodactyloidea, Tapejaridae, Thalassodrominae). Journal of Vertebrate Paleontology, 38(2): e1443273. doi:10.1080/02724634.2018.1443273
갤러리
2 장의 이미지 투푸수아라
투푸수아라투푸수아라 · 백악기 · 잡식
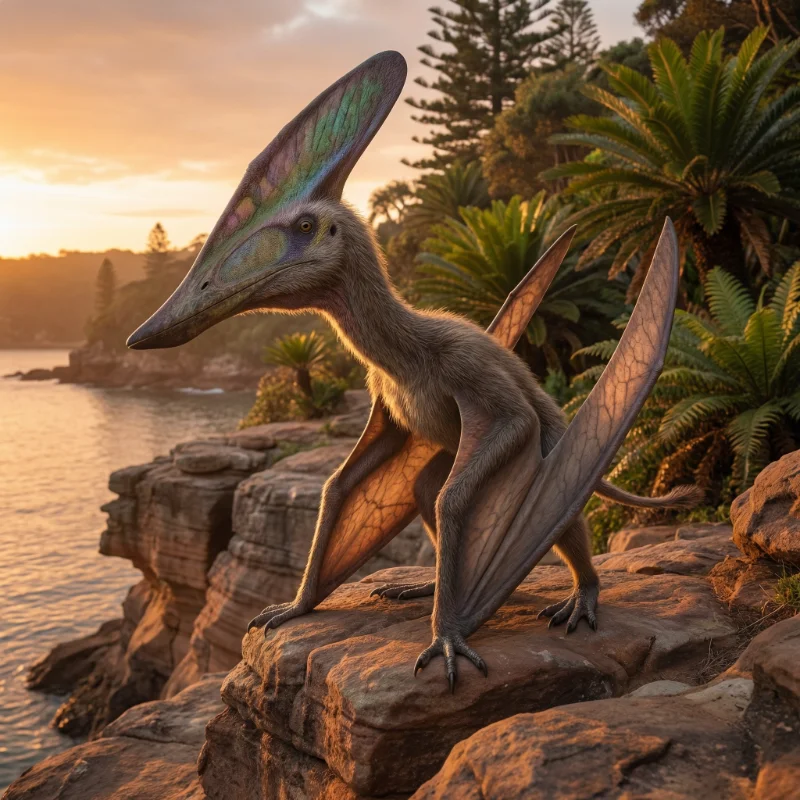 투푸수아라
투푸수아라투푸수아라 · 백악기 · 잡식







